


Sabemos que, la masa muscular de las personas desentrenadas se altera de acuerdo a la variación en el número de fibras que tiene cada músculo en particular.
Lexell et al (1988) contaron entre 393.000 y 903.000 fibras musculares esqueléticas en las secciones del vasto lateral de un grupo de 9 cadáveres de aproximadamente 19 años. Si bien existe una gran variación en el número de fibras musculares entre los individuos de este grupo, el número de las mismas en personas adultas es relativamente fijo.
Sin embargo, gracias al entrenamiento podemos incrementar las fibras de nuestros músculos.
Una persona que tiene músculos pequeños en las piernas nunca será capaz de desarrollar extremidades inferiores como las que pueden observarse en fisicoculturistas de élite, que tienen muchas fibras hipertrofiadas.
El entrenamiento con sobrecarga de alta intensidad resulta en un rápido incremento de la síntesis de proteínas fibrilares en los músculos ejercitados (Chesley et al., 1992). La respuesta llega a su pico aproximadamente luego de 24 horas de finalizada la sesión de entrenamiento, pero se mantiene elevada durante 36-48 horas (MacDougall et al., 1995, Phillips et al., 1997).
Este incremento en la síntesis es acompañado por un crecimiento proporcionalmente menor en la tasa de degradación proteica (Biolo et al., 1995, Phillips et al., 1997), tal que el efecto neto desarrolla un aumento en el balance proteico.
La suba en la tasa de síntesis de proteínas aparentemente está mediada por una traducción más eficiente del ARNm (ácido ribonucleico mensajero), ya que se produce en ausencia de cualquier cambio en el contenido total de ARN o de ARNm para proteínas contráctiles (Chesley et al., 1992, Welle et al., 1999).
Ahora veamos, con el entrenamiento de la fuerza, el incremento de la tasa sintética de proteínas se manifiesta como un aumento tanto en el área miofibrilar como en el número de miofibrillas, pero sin cambios en la densidad de empaquetamiento miofibrilar.
Los filamentos de actina y miosina se adicionan a la periferia de cada miofibrilla, aumentando el tamaño de las miofibrillas sin alterar la densidad del empaquetamiento miofibrilar o el espaciamiento de los puentes cruzados (MacDougall).
Debido a que el incremento en el área fibrilar total, en proporción, excede el incremento promedio en el área miofibrilar, es claro que además debe producirse un incremento en el número de miofibrillas.
Se cree que el incremento en el número de miofibrillas es el resultado de la “separación” longitudinal, ya que se ha mostrado que este proceso ocurre durante el crecimiento postnatal normal de animales jóvenes (Goldspink, 1970, Goldspink and Howells, 1974).
Cuando la miofibrilla alcanza un tamaño importante y una capacidad para generar tensión crítica se estima que las contracciones potentes causan lágrimas o la ruptura del tejido conectivo en los discos Z. Esto se transmite a lo largo de las miofibrillas dando como resultado dos o más “miofibrillas hijas” de igual longitud.
Nuestro objetivo hoy será revisar los procesos moleculares involucrados en el incremento del tamaño fibrilar.
¡Empecemos!
1.NUTRICIÓN Y SÍNTESIS DE PROTEÍNAS MUSCULARES
Sabemos que la masa muscular es relativamente constante a lo largo de la vida adulta, y que esto se consigue mediante el equilibrio en los procesos de síntesis y degradación proteica.
Sin embargo, con el paso de los años, este equilibrio se ve alterado y la función muscular comienza a disminuir dando como consecuencia la pérdida de masa muscular; que a su vez es resultado de la atrofia de las fibras musculares y la reducción en el número de unidades motoras,conocida como sarcopenia.
Nuestros cuerpos son capaces de mantener el equilibrio entre la síntesis y degradación proteica gracias a la ingesta de alimentos con proteínas, que resulta en hiperaminoacidemia. Así, la circulación de nutrientes anabólicos (en particular la leucina) es censada por un complejo proteico conocido como mTOR (diana de la rampamicina en mamíferos o mammalian target of rampamycin) dando por resultado un incremento considerable de la síntesis proteica a través del aumento en los procesos de iniciación y elongación de la traducción (Fujita et al., 2007).
¿Pero qué es exactamente la mTOR?El Complejo mTOR es considerado un integrador clave de múltiples estímulos positivos y negativos que pueden afectar a la masa muscular y, como dijimos antes, está implicado en el control de la iniciación de la traducción, un evento fundamental en la regulación de la síntesis proteica, mediante la fosforilación de otros componentes en la vía de señalización que modulan la unión del ARNm al complejo ribosomal 43S.
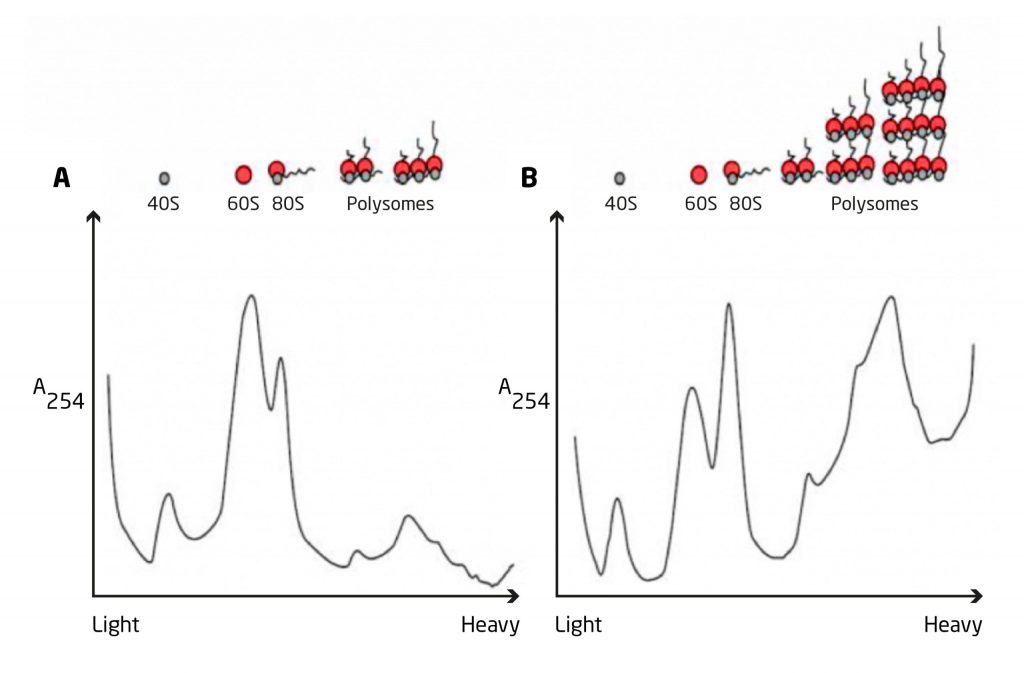
Vemos entonces que en individuos saludables, la regulación de la masa muscular depende más de las modificaciones en la síntesis de proteínas que de las modificaciones en la degradación proteica (Phillips, 2009).
Así, la tasa de síntesis proteica a nivel celular depende tanto de la eficiencia del proceso de traducción (síntesis de proteínas por unidad de ARNm) como de la capacidad de traducción (contenido ribosomal por unidad de tejido).
La síntesis de proteínas musculares ha mostrado depender de la ingesta de aminoácidos esenciales (Volpi et al., 2003), aunque la ingesta de leucina por sí sola puede actuar como señal estimulante (Anthony et al., 2002). Por esta razón, algunos autores (Bohe et al., 2003) han propuesto que existe un sensor extracelular de la concentración de aminoácidos esenciales (por ejemplo leucina); mientras que otros (Wackerhage and Rennie, 2006) sugieren que la leucina es capaz de activar directamente a la mTOR en adición de la activación de la Akt (proteína quinasa B, PKB). Estos autores mostraron que la señalización de proteína señalizadora p70S6k es independiente de la dosis de proteínas ingerida, lo que sugiere que el ejercicio ya ha estimulado completamente la maquinaria señalizadora, y que la síntesis de proteínas se vuelve esencialmente dependiente de la disponibilidad de aminoácidos.
En la actualidad todavía no se ha podido resolver la discordancia entre los eventos de señalización y la acumulación de proteínas musculares ni tampoco establecer en forma clara la manera en que los aminoácidos, solos o en combinación con ejercicio de sobrecarga, afectan la síntesis de proteínas musculares.
2. EL EJE IGF – AKT – MTOR
Vayamos por partes,
Los IGF-1 son hormonas peptídicas cuya composición es similar en el 50% de sus aminoácidos a la insulina.
A diferencia del gen de la insulina, el gen de IGF-1 codifica múltiples isoformas en múltiples tejidos. Los IGF- 1 actúan como una hormona circulante y como un factor de crecimiento local y su acción es modulada por una familia de seis proteínas de unión a IGF, así como también a través de la regulación post-transcripcional que involucra a micro ARN (miARN) a nivel tisular (Lee and Gorospe, 2010).
De esta forma, vemos que el músculo es capaz de generar un péptido diferente del canónico y maduro IGF-1 y al que se lo ha denominado IGF-1Eb o factor mecánico de crecimiento (mechano-growth factor, MGF). Hoy en día se considera que los MGF son más relevantes para el crecimiento muscular compensatorio que las isoformas sistémicas de IGF-1 (Harridge, 2007).
Es decir, según esta teoría, s los MGF ayudan en el “inicio” del proceso adaptativo post-ejercicio, resultando en un incremento de la acumulación de proteínas musculares y en la reparación del daño tisular local.
En su análisis Bamman (2007) categorizó a los sujetos en:
a) respondedores extremos(hipertrofia miofibrilar media, 58%),
b) respondedores moderados (hipertrofia miofibrilar media, 28%)
c) no respondedores (sin incremento en la hipertrofia).
Es así que, mediante la respuesta hipertrófica a un programa de entrenamiento de sobre carga de 16 semanas observó que el MGF era expresado diferencialmente en cada grupo. En los sujetos respondedores extremos registró un incremento del 126% , mientras que en los sujetos no respondedores no se registraron cambios .
Los resultados que aquí vemos, nos sugieren que el incremento transitorio inducido por el ejercicio en la expresión de MGF es una importante respuesta relacionada con la hipertrofia.
Se cree que los MGF regulan la hipertrofia de diversas maneras. Una de ellas es actuar directamente en las fibras musculares para influenciar sobre la síntesis proteica; presumiblemente ejerciendo efectos sobre la vía PI3-K – Akt –mTOR y finalmente sobre la p70S6k.
Por otra parte, existe evidencia de que los MGF pueden mantener la síntesis proteica, provocando la regulación en baja de los procesos de señalización catabólica involucrados en la degradación de proteínas. Específicamente, la evidencia sugiere que la producción local de IGF-1 suprime la localización nuclear y la actividad transcripcional de FOXO (Goodman et al., 2011).
Estas acciones combinadas ayudan a inducir una mayor acumulación de proteínas post-ejercicio. Además, los MGF parecen mediar la hipertrofia compensatoria al regular la actividad de las células satélite, ya que se ha observado que estos, están involucrados en la fase inicial de la actividad de las células satélite al ser factores críticos en la activación y proliferación de estas células (Hill et al., 2003).
Podemos afirmar entonces que, los MGF asisten a incrementar el número de mioblastos disponibles para la reparación celular post-ejercicio y facilitan la reposición del pool celular de células satélite.
3. RECEPTORES DE INSULINA Y DE IGF
Los músculos esqueléticos contienen receptores para la unión con insulina, IGF-1 y/o IGF-2. Los receptores de insulina están involucrados en la regulación metabólica, los receptores de IGF-1 están implicados en el control de crecimiento y desarrollo normal. En el tejido muscular, los receptores de IGF-1 regulan diversos aspectos de la miogénesis dirigida por las células satélites, tal como su proliferación, diferenciación y fusión.
La función de los receptores de IGF-1 es activar diversas vías de señalización intracelular; principalmente la vía de la fosfatidil inositol 3 kinasa (PI3-K) – Akt – mTOR, pero también la vía de la ERK-MAPK y la vía de la PKC. La vía de la PI3-K es considerada la vía principal a través de la cual se estimula la síntesis proteica en respuesta a los IGF-1 y en parte, en respuesta a la actividad mecánica.
Sin embargo, también se ha mostrado que la señalización vía IGF-1 no es crítica para la inducción de hipertrofia muscular, y que existen otros mecanismos para la activación de la mTOR (Spangenburg et al., 2008).
4. PROTEÍNAS AKT
Es importante tener en cuenta que, la familia de proteínas Akt consiste de tres genes que codifican las proteínas: Akt1, Akt2 y Akt3.
En los músculos esqueléticos la Akt1 está involucrada en el proceso de hipertrofia, mientras que la Akt2 desempeña un papel dominante en la absorción de glucosa estimulada por insulina.
Vimos que la activación de la Akt es suficiente para inducir hipertrofia muscular (Lai et al., 2004); sin embargo, los factores que controlan la activación de la Akt en humanos durante el ejercicio no han sido completamente dilucidados; y mucho de lo que se sabe proviene de experimentos en cultivos celulares y del rol del eje IGF-1 – Akt – mTOR en otros tejidos y tumores.
Una importante función de la Akt es que la Akt activada fosforila a la TSC2 en el complejo TSC1/TSC2. Este complejo se transloca desde el sarcolema al citosol y activa a una proteína denominada Rheb (Ras homolog enriched in brain) que l, en su forma activada, es el regulador dominante y positivo del complejo mTORC1 (complejo 1 de la mTOR).
Otra función de la Akt es que suprime la actividad de la GSK3β (glucógeno sintasa kinasa β); una enzima que ha mostrado tener un efecto inhibitorio sobre el factor 2B de iniciación de la traducción en eucariotas (elF-2B) (Mohammad- Qureshi et al., 2008) y una proteína que incrementa el proceso de traducción.
Además de estimular la hipertrofia muscular, la Akt también inhibe la inducción de la señalización de procesos asociados con la atrofia muscular a través FOXO (Stitt et al., 2004).
En este sentido, se sugiere que el eje IGF-1 – Akt – mTOR no es solo instrumental en incrementar la síntesis proteica sino que también es capaz de contrarrestar la señalización atrófica que se produce en ciertos estados patológicos (sepsis, cachexia, altas dosis externas de glucocorticoides, diabetes, etc.) a través de factores como el factor de necrosis tumoral tipo alfa (TNF-α) o interleucina 1 (IL-1).
La atrofia muscular en estas condiciones depende parcialmente de la inducción de ubiquitina-ligasas E3, tales como MuRF-1 y MAFbx (también denominada atrogina-1) (Sacheck et al., 2004) que ubiquitina proteínas musculares estructurales tales como las cadenas pesadas de miosina. Estas proteínas son así objetivo de la degradación por proteosomas.
En este proceso los factores de transcripción FOXO1/3 (forkhead box O1/O3) demostraron ser importantes para el control transcripcional de MuRF-1 y MAFbx. Y los experimentos con ejercicios de sobrecarga en humanos mostraron que este tipo de ejercicio es capaz de atenuar la expresión genética de MuRF-1 y MAFbx (Mascher et al., 2008).
5. MTOR Y PROTEÍNAS ASOCIADAS
El complejo mTOR es una proteína regulatoria clave para procesos celulares tales como la proliferación, el crecimiento y la diferenciación celular, la síntesis proteica, la organización citoesquelética, la homeostasis energética y el metabolismo de los sustratos (Rivas et al., 2009).
El principal efector de la mTOR, corriente abajo en la vía de señalización, es la proteína quinasa ribosomal S6 β-1 (p70s6k). La mTOR es tanto un componente integral de la cascada de señalización de la insulina como un sensor de nutrientes independiente de la insulina (Hawley and Lessard, 2008, Krebs et al., 2007). Además,es un mediador clave de las adaptaciones inducidas por el entrenamiento con sobrecarga en los músculos esqueléticos (Coffey and Hawley, 2007).
Asimismo, esta proteína existe como dos complejos que son estructural y funcionalmente diferentes, la mTORC1 y la mTORC2 (Figura 2).
La mTORC1 está asociada con diferentes proteínas, incluida la proteína raptor mientras que la mTORC1 tiene a la p70s6K como objetivo primario de fosforilación y mejora el proceso de traducción mediante la inhibición de la proteína de unión al factor de iniciación de la traducción en ecuariotas 4E (4E-BP1).En consecuencia, la mTORC1 es una vía de señalización crítica para el control de la síntesis de proteínas en respuesta a la nutrición y el ejercicio (Drummond et al., 2009). Estos autores señalan que la mTORC1 es un nodo clave en la señalización intracelular muscular que responde a señales de la insulina, al estatus energético, a la concentración de aminoácidos y otros nutrientes, y también a factores de crecimiento.
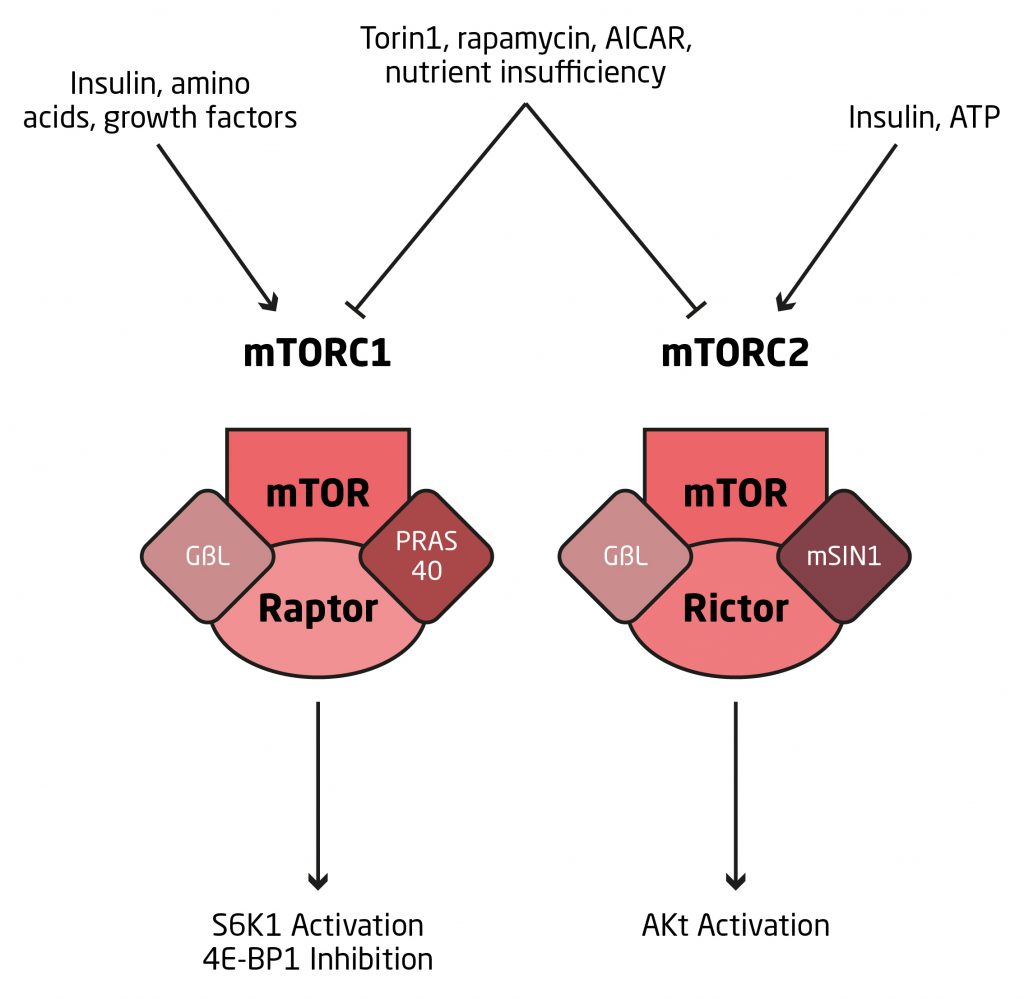
Por otro lado, la mTORC2, asociada entre otras proteínas a la proteína rictor, es un regulador positivo de la absorción de glucosa estimulada por la insulina, a través de la retroalimentación de la Akt; pero no está involucrada en la señalización iniciada por la contracción (Kumar et al., 2008).
Como ya mencionamos, la mTOR está positivamente regulada por factores de crecimiento (IGF-1) y por la insulina a través de la vía PI3-K – Akt – TSC1/TSC2 – Rheb (Figura 3). Además, se ha establecido que la disponibilidad de aminoácidos (específicamente leucina) es un importante regulador de la actividad de la mTOR durante protocolos de entrenamiento con sobrecarga en humanos (Dreyer et al., 2008).
Sugerimos además, que las proteínas “Rag” (una familia de cuatro GTPasas) pueden estimular la actividad de la mTOR posiblemente al colocar la MTOR con su activador Rheb (Kim et al., 2008).
Uno de los aspectos más importantes de la mTOR, que fue observado por primera vez por Bodine et al. (2001b), es la fuerte asociación entre la carga mecánica y la activación de la mTOR.
La carga mecánica es suficiente para activar la mTOR y esta puede ocurrir independientemente de la estimulación de la vía PI3-K; lo cual implica una contribución del citoesqueleto celular a la activación de la mTOR con la carga mecánica.
En este contexto, las quinasas de adhesión focal (FAK) han mostrado desempeñar un rol importante en la señalización de la síntesis de proteínas musculares reguladas mecánicamente y pudimos ver que la mTOR puede ser activada independientemente de la vía PI3- K/Akt mediante la intervención de la fosfolipasa D (PLD) y su segundo mensajero el ácido fosfatidico (PA) (Hornberger et al., 2006).
Por otro lado, establecimos que la mTOR es sensible al estatus energético celular a través de la señal enviada por la vía de la AMPK. Al parecer, la fosforilación de la TSC2 dependiente de la AMPK deriva en la inhibición de la actividad de la mTOR en condiciones de baja energía (Inoki et al., 2006).
Aquí, ahora la evidencia sugiere que la AMPK es capaz de fosforilar directamente a la proteína raptor unida a la mTORC1, inhibiendo así la actividad de ésta. Asimismo, se ha identificado que las proteínas REDD1 y REDD2 son factores que regulan negativamente a la mTOR (Thomson et al., 2008). Se ha sugerido, también, que las REDD pueden mediar la comunicación cruzada entre los procesos de censado de oxígeno y el crecimiento.
6. PROTEÍNAS REDD1 Y REED2
Diversas líneas de investigación sugieren que la REDD1 y la REDD2 pueden regular negativamente a la mTOR afectando la activación de la Akt (corriente abajo en la vía de señalización) y a la TSC2 (corriente arriba en la vía de señalización) bajo diversas condiciones de estrés tales como hipoxia, intoxicación con alcohol y altas dosis de glucocorticoides (Miyazaki et al., 2010).
Además, observamos que con el entrenamiento de sobrecarga de baja intensidad la expresión de ARNm de REDD1 se mantuvo significativamente reducida incluso 3 hs después del ejercicio, mientras que la expresión de ARNm de REDD2 se mantuvo reducida hasta 6 hs después de un único bloque de entrenamiento con sobrecarga de alta intensidad combinado con la ingesta de aminoácidos esenciales (Drummond et al., 2009).
Esto sugiere que la vía de las REDD son relevantes para el control del tamaño muscular inducido por la contracción en humanos, a través del control negativo de la mTOR.
En definitiva tras este detallado y completo análisis podemos ver que el eje IGF-1 – Akt – mTOR es el principal integrador de señales relacionadas al mantenimiento e incremento inducido por el entrenamiento del tejido muscular de adultos.
Diversas vías de señalización controladas por retroalimentación, como las citocinas, el estímulo mecánico y el estatus nutricional de las células musculares, integran la información modificando diversos eventos en múltiples sitios que controlan los procesos de traducción e influencian la masa muscular.
Además, se produce una importante comunicación entre estas vías y las vías de señalización relacionadas con el estatus energético y el metabolismo.
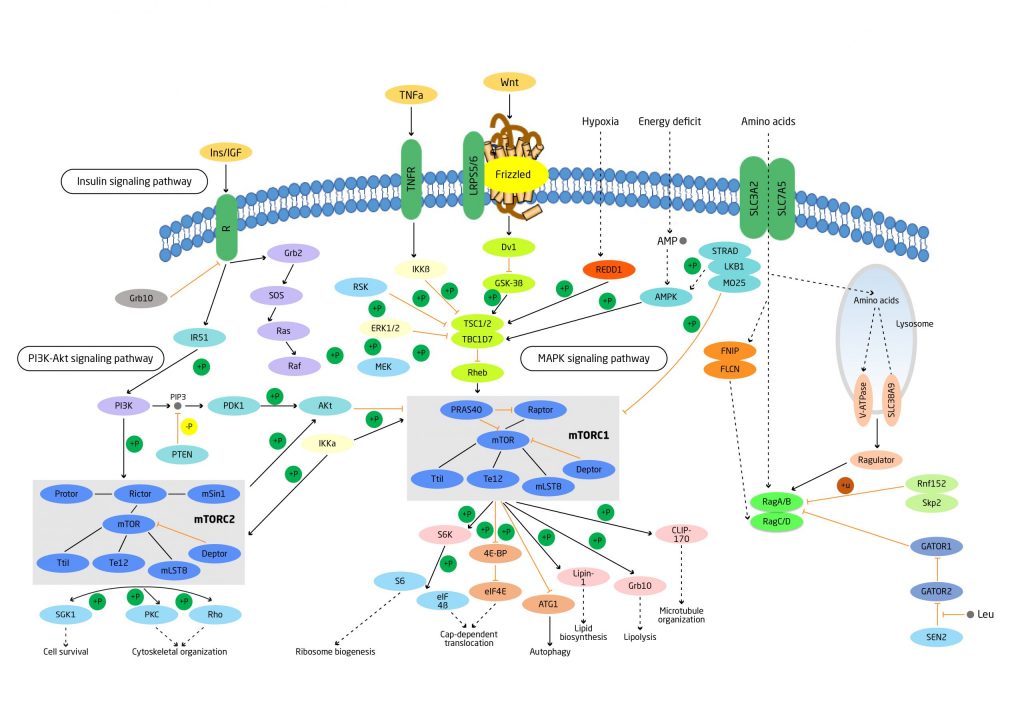
La vía del complejo mTORC1 puede ser activado por hormonas, factores de crecimiento y nutrientes. Diferentes quinasas modulan la actividad de la mTORC1 convergiendo en el complejo TSC1/TSC2. Bajos niveles de glucosas, ATP y oxígeno, así como el daño del ADN pueden derivar a en la activación de la AMPK que inhibe a la mTORC1 activando el complejo TSC1/TSC2 o directamente fosforilando la proteína raptor componente de la mTORC1.
La activación de la mTORC1 a su vez deriva en la activación de la p70s6k y la proteína de unión al factor de iniciación de la traducción en eucariotas (4E-BP1) que modulan diversas funciones celulares.
El complejo mTORC2 puede ser activado por insulina y factores de crecimiento a través de la vía de la PI3-K. A su vez, la mTORC2 activa la Akt facilitando así la activación de la mTORC1. Reproducido de Haissaguerre et al. (2014).
¿Querés aprender más? Elegí alguno de nuestros cursos y pagalo con un 20% de descuento si ya sos miembro. ¿Aún no sos miembro? ¡Dale clic acá y sumate!





